Jaa:



Liitotyypit ja niiden ominaisuudet
solujen liitokset ne ovat kosketussiltoja, jotka ovat vierekkäisten solujen välisten sytoplasmisten kalvojen välillä tai solun ja matriisin välillä. Nivelet riippuvat tutkitun kudoksen tyypistä ja korostavat epiteelisolujen, lihasten ja hermosolujen välisiä yhteyksiä.
Soluissa on molekyylejä, jotka liittyvät niiden väliseen tarttuvuuteen. Tarvitaan kuitenkin lisäelementtejä, jotka lisäävät kudosten sitoutumisen vakautta. Tämä saavutetaan solujen liitoksilla.
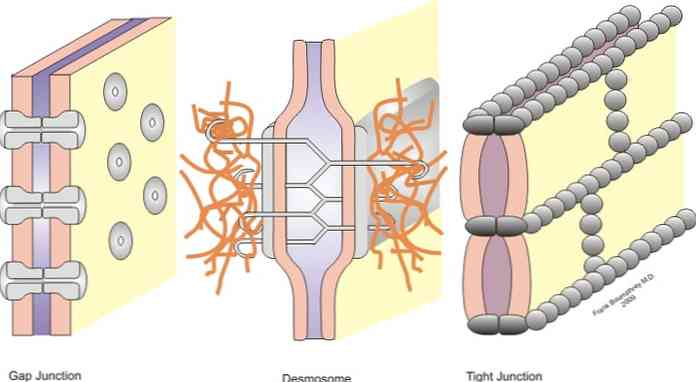
Liitokset luokitellaan symmetrisiin liitoksiin (kapeat liitokset, hihnan desmosomit ja rakonivelet) ja epäsymmetriset nivelet (hemidesmosomit).
Kapeat liitokset, hihnan desmosomit, pistemäiset desemomeot ja hemidesmosomit ovat liitoksia, jotka mahdollistavat ankkuroinnin; kun taas raon liitokset käyttäytyvät naapurisolujen välisen liiton siltojena, mikä mahdollistaa solujen vaihtamisen sytoplasmien välillä.
Liuottimien, veden ja ionien liikkuminen tapahtuu yksittäisten solukomponenttien läpi ja niiden välillä. Täten on olemassa transcellulaarinen reitti, jota ohjaa sarja putkia ja kuljettimia. Toisin kuin parasellulaarinen polku, jota säätelevät solujen väliset yhteydet - eli solujen liitokset.
Kasveissa löytyy solukkoliittymiä, jotka muistuttavat raon liitoksia, joita kutsutaan plasmodmeiksi. Vaikka ne eroavat toisistaan, se on sama.
Lääketieteellisestä näkökulmasta tietyt solujen risteyksien puutteet johtavat saavutettuihin tai perinnöllisiin sairauksiin, jotka ovat aiheutuneet epiteelisulun vaurioitumisesta.
indeksi
- 1 Ominaisuudet
- 2 tyyppiä
- 2.1 - Kapeat yhteydet
- 2.2 - Raon tai aukon liitokset
- 2.3 - Ankkurointi tai kiinnittyminen
- 2.4 -Hemidesmosoomit
- 3 Solukkoliittymät kasveissa
- 4 Lääketieteellinen näkökulma
- 5 Viitteet
piirteet
Elävät organismit koostuvat erillisistä ja monipuolisista rakenteista, joita kutsutaan soluiksi. Nämä rajataan plasmamembraanilla, joka pitää ne erillään solunulkoisesta ympäristöstä.
Vaikka ne ovat elävien olentojen komponentteja, ne eivät kuitenkaan muistuta tiiliä, koska ne eivät ole erillään toisistaan.
Solut ovat elementtejä, jotka ovat yhteydessä toisiinsa ja solunulkoiseen ympäristöön. Siksi solujen täytyy muodostaa kudoksia ja kommunikoida, kun kalvo pysyy ehjänä.
Tämä ongelma ratkaistaan epiteelissä esiintyvien soluliittymien läsnäolon ansiosta. Nämä liitokset muodostetaan kahden vierekkäisen solun väliin luokiteltuna kunkin symmetristen ja epäsymmetristen liitosten funktion mukaan.
Epäsymmetriset yhteydet kuuluvat hemidesmosomeihin, ja symmetrisissä yhteyksissä läheiset yhteydet, hihnan desmosoma, desmosomit ja rako-yhteydet. Seuraavaksi kuvataan yksityiskohtaisesti kutakin ammattiliittoa.
tyyppi
-Kapeat risteykset

Kapeat liitokset, jotka tunnetaan myös kirjallisuudessa okklusiivisina liitoksina, ovat naapurisolujen solukalvojen sektoreita, jotka ovat läheisesti sidoksissa - kuten nimi "kapea liitos" osoittaa.
Keskimääräisissä olosuhteissa solut erotetaan etäisyydellä 10 - 20 nm. Kuitenkin kapeiden liitosten tapauksessa tämä etäisyys vähenee merkittävästi ja molempien solujen kalvot koskettavat tai jopa sulautuvat.
Tyypillinen kapea liitos sijaitsee naapurisolujen sivuseinämien vä- lillä minimietäisyydellä niiden apikaalisista pinnoista.
Epiteelis kudoksessa kaikki solut muodostavat tämän tyyppisiä liitoksia, jotta ne pysyisivät yhtenäisinä. Tässä vuorovaikutuksessa solut sijaitsevat muodostaen renkaan muistuttavan kuvion. Nämä liitot kattavat koko kehän.
Proteiinit osallistuvat tiiviisiin risteyksiin
Ocludina ja Claudina
Läheisen kosketuksen alueet ympäröivät koko solun pintaa. Nämä alueet muodostavat transmembraaniproteiinien, joita kutsutaan okklusiineiksi ja claudiiniksi, anastomoottisia kontaktiliuskoja. Termi anastomosis viittaa tiettyjen anatomisten elementtien yhteenliittämiseen.
Nämä kaksi proteiinia kuuluvat tetraespaniinien ryhmään. Niille on tunnusomaista, että niillä on neljä transmembraanidomeeniä, kaksi ulkoista silmukkaa ja kaksi suhteellisen lyhyttä sytoplasmista häntä.
On havaittu, että occludin on vuorovaikutuksessa neljän muun proteiinimolekyylin kanssa, joita kutsutaan okklonaaliksi ja jotka on lyhennetty ZO: ksi. Tämä viimeinen ryhmä sisältää proteiinit ZO1, ZO2, ZO3 ja afuna.
Toisaalta Claudin on 16 proteiinin perhe, joka muodostaa joukon lineaarisia fibrilejä kapeissa risteyksissä, mikä sallii tämän liiton ottaa vastaan "esteen" roolin parasellulaarisessa reitissä..
Nectinas ja JAM
Nektiinit ja liittojen adheesiomolekyylit (lyhennettynä lyhenteellä englanniksi JAM) näkyvät myös kapeissa risteyksissä. Nämä kaksi molekyyliä löydetään homodimeereinä solunsisäisessä tilassa.
Nektiinit on liitetty aktiinifilamentteihin afadiiniproteiinin avulla. Jälkimmäinen näyttää olevan elintärkeä, koska jyrsijöiden afadiinia koodaavan geenin deleetioissa ne johtavat alkion kuolemaan.
Kapean risteyksen toiminnot
Tämäntyyppiset solu-solu-risteykset suorittavat kaksi olennaista toimintoa. Ensimmäinen on määrittää epiteelin solujen napaisuus, erottaen apikaalisen domeenin basolateraalisesta ja estää lipidien, proteiinien ja muiden biomolekyylien kohtuuttoman diffuusion..
Kuten mainitsimme määritelmässä, epiteelin solut on ryhmitelty renkaaseen. Tämä rakenne erottaa solun apikaalisen pinnan sivuttaisista ja peruspohjoista, mikä muodostaa erottelun verkkotunnusten välillä.
Tätä erottelua pidetään yhtenä tärkeimmistä käsitteistä epiteelin fysiologian tutkimuksessa.
Toiseksi, tiukat liitokset estävät aineiden vapaan kulun epiteelisolujen kerroksen läpi, mikä muuttuu esteeksi paratsellulaariselle reitille..
-Raon tai aukon liitokset

Raon tai raon liitokset löytyvät alueista, joilla ei ole rajoittavia sytoplasmakalvoja naapurisolujen välillä. Jaetussa risteyksessä solujen sytoplasmat on kytketty ja fyysinen yhteys muodostuu, kun pienten molekyylien kulku voi tapahtua.
Tämä nivelluokka löytyy käytännöllisesti katsoen kaikista epiteeleistä ja muista kudostyypeistä, joissa ne käyttävät melko vaihtelevia tarkoituksia..
Esimerkiksi useissa kudoksissa raon liitokset voivat avautua tai sulkea vasteena solunulkoisille signaaleille, kuten neurotransmitterin dopamiinin tapauksessa. Tämän molekyylin läsnäolo vähentää verkkokalvon luokan neuronien välistä viestintää vasteena valon lisääntyneeseen intensiteettiin.
Proteiinit, jotka osallistuvat pilkkoutumisliitoksiin
Slit-liitokset muodostuvat proteiineista, joita kutsutaan konnexiineiksi. Siten "conexón" saadaan kuuden connexin-monomeenin liitosta. Tämä rakenne on ontto sylinteri, joka kulkee sytoplasmisen kalvon läpi.
Liitännät on järjestetty siten, että vierekkäisten solujen sytoplasmien välille muodostuu kanava. Lisäksi connexonit yleensä aggregaatit ja muodostavat eräänlaiset levyt.
Raonivelien toiminnot
Näiden sidosten muodostumisen ansiosta tiettyjen molekyylien liikkuminen naapurisolujen välillä voi tapahtua. Kuljetettavan molekyylin koko on ratkaiseva, optimaalinen halkaisija on 1,2, kuten kalsiumionit ja syklinen adenosiinimonofosfaatti.
Erityisesti epäorgaanisia ioneja ja vesiliukoisia molekyylejä voidaan siirtää solun sytoplasmasta viereiseen sytoplasmaan.
Kalsiumpitoisuudet ovat ratkaisevassa asemassa tässä kanavassa. Kun kalsiumpitoisuus kasvaa, aksiaaliset kanavat pyrkivät sulkeutumaan.
Tällä tavoin rakojen liitokset osallistuvat aktiivisesti solujen välisen sähköisen ja kemiallisen kytkennän prosessiin, kuten tapahtuu sydänlihassoluissa, jotka vastaavat sähköimpulssien lähettämisestä.
-Ankkurointi tai liitännät

Kapeiden liitosten alapuolella löydämme ankkurointiliitokset. Yleensä nämä sijaitsevat epiteelin apikaalisen pinnan läheisyydessä. Tässä ryhmässä voimme erottaa kolme pääryhmää, vyöhykkeellä olevat zonula-adhereenit tai desmosoomit, makulan adhereenit tai desmosomipisteet ja desmosome.
Tämäntyyppisissä liitoksissa vierekkäiset solukalvot, jotka on liitetty vyöhykkeisiin ja adhereenimakuloihin, erotetaan suhteellisen leveällä solumuovilla - jos verrataan niitä pienimpään tilaan, joka on olemassa kapeissa risteyksissä..
Intercellulaarisessa tilassa on proteiineja, jotka kuuluvat kadheriinien, desmogleiinien ja desmokoliinien perheeseen, jotka on liitetty sytoplasmisiin levyihin, joissa on muita desmoplaquina-, placoglobina- ja placofilina-proteiineja..
Kiinnityspisteiden luokitus
Zonula adherens
Kuten kapeissa liitoksissa, ankkurointiliitoksissa havaitsemme myös järjestelyn muodon renkaan tai hihnan muodossa. Zonula adherens liittyy aktinimikrofilmiin kahden proteiinin - kadheriinien ja kateniinien - vuorovaikutuksen kautta..
Macula adherens
Joissakin tapauksissa tämä rakenne tunnetaan yksinkertaisesti desmosomana, se on punkkiliitos, joka liittyy keratiinista muodostettuihin välifilamentteihin. Tässä yhteydessä mainitut keratiinirakenteet ovat nimeltään "tonofilimanetos". Filamentit ulottuvat epiteelisoluissa yhdestä pisteestä toiseen.
Point desmosomas
Nämä aikaansaavat voimakkuuden ja jäykkyyden epiteelisoluihin. Näin ollen uskotaan, että sen päätehtävä liittyy vierekkäisten solujen vahvistamiseen ja stabilointiin.
Desmosomeja voidaan verrata eräänlaisiin niitteihin tai hitseihin, koska ne muistuttavat erillisiä pieniä täpliä ja ei jatkuvia nauhoja.
Tämän tyyppiset nivelet löytyvät interkaloiduista levyistä, jotka yhdistävät sydänlihaksen sydänlihakset ja aivojen ja selkäytimen ulkopintaa peittävät aivokalvot..
-hemidesmosomin

Hemidesmosomit kuuluvat epäsymmetristen risteyksien luokkaan. Näiden rakenteiden tehtävänä on ankkuroida epiteelisolun basaalidomeeni taustalla olevan peruslaminan kanssa.
Termiä hemidesmosome käytetään, koska tämä rakenne näyttää kirjaimellisesti "keskisuurelta" desmosomalta. Biokemiallisen koostumuksen näkökulmasta molemmat ammattiliitot ovat kuitenkin täysin erilaisia.
On tärkeää selventää, että desmosomit ovat vastuussa naapurisolun kiinnittymisestä toiseen, kun taas hemidesmosomin tehtävänä on liittyä soluun basaalilevyn kanssa.
Toisin kuin makulan adhereenit tai desmosomaatit, hemidesmosomeilla on erilainen rakenne, joka koostuu: sytoplasmisesta lamiinasta, joka liittyy välituotteisiin ja ulkoisten kalvojen levyyn, joka on vastuussa hemidesmosomin liittämisestä basaalilevyyn käyttämällä ankkurifilamentti.
Yksi hemidesmosomien tehtävistä on lisätä epiteelisien kudosten yleistä stabiilisuutta, johtuen basaalilaminaatin komponentteihin kiinnitetyn sytoskeletonin välifilamenteista..
Solujen liitokset kasveissa
Kasvien valtakunnasta puuttuu useimmat edellä kuvatut solukohdat, lukuun ottamatta funktionaalista vastakappaletta, joka muistuttaa raon liitoksia.
Kasveissa vierekkäisten solujen sytoplasmat yhdistetään reiteillä tai kanavilla, joita kutsutaan plasmodesmeiksi.
Tämä rakenne luo jatkuvuuden yhdestä kasvisolusta seuraavaan. Vaikka se eroaa rakenteellisesti rakojen liitoksista, niillä on hyvin samanlaiset roolit, jolloin pienet ionit ja molekyylit kulkevat.
Lääketieteellinen näkökulma
Lääketieteen näkökulmasta soluliitot ovat tärkeä asia. On havaittu, että geeneissä olevat mutaatiot, jotka koodittavat liittymiin osallistuvia proteiineja, muunnetaan kliinisiksi patologioiksi.
Esimerkiksi jos geenissä on tietty mutaatio, joka koodaa tiettyä claudin-tyyppiä (yksi proteiineista, jotka välittävät vuorovaikutusta kapea-tyyppisissä liitoksissa), se aiheuttaa ihmisille harvinaisen sairauden..
Tämä on munuaisten magnesiumin häviön oireyhtymä, ja oireet sisältävät pienen määrän magnesiumia ja kohtauksia.
Lisäksi on havaittu, että proteiinin nektiiniä 1 koodaavan geenin mutaatio on vastuussa kurkunvaimennuksen oireyhtymästä tai halkeamiskielestä. Tätä tilannetta pidetään yhtenä vastasyntyneiden yleisimmistä epämuodostumista.
Nektiini 1 -geenin mutaatiot ovat myös liittyneet toiseen sairauteen, jota kutsutaan ektodermaaliseksi dysplasiaksi, joka vaikuttaa ihmisen ihoon, hiuksiin, kynsiin ja hampaisiin..
Foliar pemphigus on ihon bulloosinen patologia, joka määritetään desmogleiini 1: n vastaisilla auto-vasta-aineilla, joka on keskeinen tekijä, joka on vastuussa epidermiksen yhteenkuuluvuuden ylläpitämisestä.
viittaukset
- Alberts, B., Bray, D., Hopkin, K., Johnson, A.D., Lewis, J., Raff, M., ... & Walter, P. (2015). Oleellinen solubiologia. Garland Science.
- Cooper, G. M., ja Hausman, R. E. (2000). Solu: Molekyylinen lähestymistapa. Sinauer Associates.
- Curtis, H., & Barnes, N. S. (1994). Kutsu biologiaan. Macmillan.
- Hill, R. W., Wyse, G. A., Anderson, M., ja Anderson, M. (2004). Eläinten fysiologia. Sinauer Associates.
- Karp, G. (2009). Solu- ja molekyylibiologia: käsitteet ja kokeilut. John Wiley & Sons.
- Kierszenbaum, A., & Tres, L. (2016). Histologia ja solubiologia: johdanto patologiaan. Elsevier Brasilia.
- Lodish, H., Berk, A., Darnell, J.E., Kaiser, C. A., Krieger, M., Scott, M.P., ... & Matsudaira, P. (2008). Molekyylisolubiologia. Macmillan.
- Voet, D., & Voet, J. G. (2006). biokemia. Ed. Panamericana Medical.