Jaa:



Topoisomeraasien ominaisuudet, toiminnot, tyypit ja estäjät
topoisomeraaseihin ovat erään tyyppisiä isomeraasientsyymejä, jotka muuttavat deoksiribonukleiinihapon (DNA) topologiaa ja tuottavat sekä sen purkamisen käämityksessään että superkäämiään.
Näillä entsyymeillä on erityinen rooli DNA: n vääntöjännityksen lievittämisessä niin, että tärkeitä prosesseja, kuten sen replikaatiota, DNA: n transkriptiota messenger-ribonukleiinihapossa (mRNA) ja DNA: n rekombinaatiota voi esiintyä..

Topoisomeraasientsyymejä on läsnä sekä eukaryoottisissa että prokaryoottisissa soluissa. Tutkijat Watson ja Crick ennustivat sen olemassaolon arvioidessaan rajoituksia, joita DNA-rakenne esitti salliakseen sen tietoihin pääsyn (tallennetaan sen nukleotidisekvenssiin).
Topoisomeraasien toimintojen ymmärtämiseksi on katsottava, että DNA: lla on stabiili kaksoiskierteen rakenne, jossa ketjut on rullattu toisen päälle.
Nämä lineaariset ketjut muodostuvat 2-deoksiriboosista, joka on liitetty fosfodiesterisidoksiin 5'-3 ', ja typpipohjaisista emäksistä niiden sisätiloissa, kuten kierreportaat..

Topologinen tutkimus DNA-molekyylien on osoittanut, että nämä voivat omaksua useita konformaatioita riippuvainen sen vääntövoima: alkaen lepotilassa eri valtioiden jolloin käämin tiivistys.
DNA-molekyylejä, joilla on erilaiset konformaatiot, kutsutaan topoisomeereiksi. Näin ollen voimme päätellä, että topoisomeraasit I ja II voivat lisätä tai pienentää DNA-molekyylien vääntöjännitystä muodostaen niiden erilaiset topoisomeerit.
Mahdollisten DNA-topoisomeerien joukossa yleisin konformaatio on superkääminen, joka on hyvin kompakti. Topoisomeraasien täytyy myös irrottaa DNA: n kaksoiskierukka useiden molekyyliprosessien aikana.
indeksi
- 1 Ominaisuudet
- 1.1 Yleisen toiminnan mekanismi
- 1.2 Topoisomeraasit ja solusykli
- 2 Toiminnot
- 2.1 Geneettisen materiaalin kompakti varastointi
- 2.2 Geneettisten tietojen saatavuus
- 2.3 Geenin ilmentymisen säätely
- 2.4 Topoisomeraasi II: n erityispiirteet
- 3 Topoisomeraasien tyypit
- 3.1-topoisomeraasit, tyyppi I
- 3.2 -Topoisomeraasit, tyyppi II
- 3.3 - Ihmisen topoisomeraasit
- 4 Topoisomeraasin estäjät
- 4.1-Topoisomeraasit kemiallisen hyökkäyksen kohteena
- 4.2 -Kääntymisen tyypit
- 4.3 - Topoisomeraasi-inhibiittorit
- 5 Viitteet
piirteet
Yleisen toiminnan mekanismi
Jotkut topoisomeraasit voivat rentoutua vain negatiivisia DNA-supersoluja, tai molempia DNA-supersoluja: positiivisia ja negatiivisia.
Jos pyöreä kaksijuosteista DNA kiertyy pituusakselinsa ympäri ja vasenkätisen puolestaan tapahtuu (suuntaan myötäpäivään), sanotaan olevan negatiivisesti superkierteistä. Jos puolestaan on myötäpäivään (vastapäivään myötäpäivään), se on positiivisesti superkierteistä.

Periaatteessa topoisomeraasit voivat:
-Helpotetaan DNA-juosteen kulkua vastakkaiseen säikeeseen leikatun leikkauksen avulla (topoisomeraasi tyyppi I).
-Helpotetaan täydellisen kaksoiskierroksen kulkua halkeaman läpi itsestään tai jakautumalla toiseen eri kaksoiskierteeseen (topoisomeraasi tyyppi II).
Yhteenvetona voidaan todeta, että topoisomeraasit toimivat fosfodiesterisidosten katkaisemisen kautta yhdessä tai molemmissa säikeissä, jotka muodostavat DNA: n. Sitten muokkaa kaksoiskierteen (topoisomeraasi I) tai kahden kaksoisheliksin (topoisomeraasi II) kierreiden käämitystilaa, jotta lopulta palataan sitomaan tai sitomaan leikatut päät.
Topoisomeraasit ja solusykli
Vaikka topoisomeraasi I on entsyymi, jolla on suurempi aktiivisuus S-vaiheen aikana (DNA-synteesi), sen ei katsota olevan riippuvainen solusyklin vaiheesta.
Vaikka topoisomeraasi II -aktiivisuus on aktiivisempi solun kasvun logaritmisessa vaiheessa ja nopeasti kasvavien kasvainten soluissa.
tehtävät
Topoisomeraaseja koodaavien geenien muutos on soluille tappava, mikä osoittaa näiden entsyymien merkityksen. Prosesseista, joissa topoisomeraasit osallistuvat, kuuluvat:
Geneettisen materiaalin kompakti varastointi
Topoisomeraasit varastoinnin helpottamiseksi geneettisen informaation tiiviisti, koska ne tuottavat kiemura ja DNA superkierteisyyden jolloin suuri määrä tietoa on suhteellisen pieni tilavuus.
Pääsy geneettiseen tietoon
Jos oli topoisomeraaseja ja sen ainutlaatuiset ominaisuudet, olisi mahdotonta saada tallennettua tietoa DNA: ta. Tämä johtuu siitä, topoisomeraasit määräajoin vapauttamaan vääntöjännitys syntyy kahden DNA-kierteen, kun taas purkautuminen prosessit replikaatiota, transkriptiota ja rekombinaatio.

Jos näiden prosessien aikana syntyneestä vääntymisestä johtuvaa jännitystä ei vapauteta, se voi aiheuttaa viallisen geeniekspressiota, pyöreän DNA: n tai kromosomin keskeytymisen, jopa tuottaa solukuoleman.
Geenin ilmentymisen säätely
DNA-molekyylin konformaatiomuutokset (kolmiulotteisessa rakenteessa) altistavat tietyt alueet ulkopuolelle, jotka voivat olla vuorovaikutuksessa DNA: ta sitovien proteiinien kanssa. Näillä proteiineilla on säätelevä funktio geenien ilmentymisestä (positiivinen tai negatiivinen).
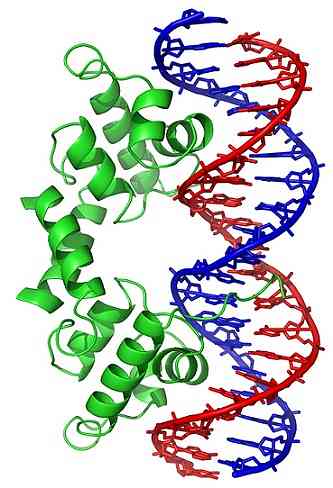
Siten topoisomeraasien vaikutuksesta muodostettu DNA-käämityksen tila vaikuttaa geeniekspression säätelyyn.
Topoisomeraasin II erityispiirteet
Topoisomeraasi II: n on tarpeen kokoonpanoon kromatidien, tiivistyminen ja decondensation kromosomit ja erottelu tytär DNA-molekyylien mitoosin aikana.
Tämä entsyymi on myös rakenteellinen proteiini ja yksi solun ytimen matriisin tärkeimmistä aineosista interfaasin aikana..
Topoisomeraasien tyypit
Topoisomeraaseja on kaksi päätyyppiä riippuen siitä, voivatko ne pilkkoa yhtä tai kahta DNA-säikettä.
-Tyypin I topoisomeraasit
monomeeriset
Tyypin I topoisomeraasit ovat monomeerejä, jotka lieventävät negatiivisia ja positiivisia supersäikeitä, jotka tuotetaan haarukan liikkumisen aikana transkription aikana, ja replikaation ja geenin rekombinaation prosessien aikana.
Tyypin I topoisomeraasit voidaan jakaa tyypin 1A ja tyypin 1B mukaan. Viimeksi mainitut ovat ihmisissä havaittuja, ja ne ovat vastuussa supersuojennetun DNA: n rentoutumisesta.
Tyrosiini sen aktiivisessa paikassa
Topoisomeraasi 1B (Top1B) koostuu 765 aminohaposta, jotka on jaettu neljään spesifiseen domeeniin. Yhdessä näistä domeeneista on erittäin säilynyt alue, joka sisältää aktiivisen kohdan tyrosiinin (Tyr7233) kanssa. Kaikki topoisomeraasit, jotka ovat aktiivisessa kohdassa, ovat tyrosiini, jolla on olennainen rooli koko katalyyttisessä prosessissa.
Toimintamekanismi
Aktiivisesta kohdasta peräisin oleva tyrosiini muodostaa kovalenttisen sidoksen DNA-juosteen 3'-fosfaattipäähän, leikkaa sen pois ja pitää sen kiinnittyneenä entsyymiin samalla, kun se kulkee toisen DNA-juosteen ulokkeen läpi.
Kulkua muiden DNA-juosteen läpi katkaistun juosteen konformationaalisen muutoksen entsyymin, joka tuottaa aukko kaksoiskierteen DNA saavutetaan.
Sitten topoisomeraasi I palaa alkuperäiseen konformaatioonsa ja sitoutuu uudelleen leikattuihin päihin. Tämä tapahtuu prosessilla, joka on käänteinen DNA-ketjun rikkoutumiseen entsyymin katalyyttisessä kohdassa. Lopuksi topoisomeraasi vapauttaa DNA-juosteen.
DNA-ligointinopeus on korkeampi kuin leikkausnopeus, joka takaa molekyylin stabiilisuuden ja genomin eheyden.
Yhteenvetona voidaan todeta, että tyypin I topoisomeraasi katalysoi:
- Lohkon poisto.
- Toisen juosteen kulku jakauman kautta.
- Jaettujen päiden ligaatio.
-Tyypin II topoisomeraasit
dimeerinen
Tyypin II topoisomeraasit ovat dimeerisiä entsyymejä, jotka pilkkovat molempia DNA-säikeitä, mikä rentouttaa transkription ja muiden soluprosessien aikana syntyneet superkuvat..
Mg-riippuvaiset++ ja ATP
Nämä entsyymit tarvitsevat magnesiumia (Mg++) ja tarvitsevat myös energiaa, joka on peräisin ATP-trifosfaattiyhteyden hajoamisesta, jota he käyttävät ATPaasin ansiosta.
Kaksi aktiivista kohtaa, joissa on tyrosiinia
Ihmisen topoisomeraasit II ovat hyvin samankaltaisia kuin hiivan (Saccharomyces cerevisiae), joka koostuu kahdesta monomeeristä (A ja B). Kussakin monomeerissä on ATPaasi-domeeni ja alafragmentissa tyrosiiniaktiivinen kohta 782, johon DNA voi sitoutua. Siksi kaksi DNA-säikettä voidaan liittää topoisomeraasiin II.
Toimintamekanismi
Topoisomeraasi II: n vaikutusmekanismi on sama kuin topoisomeraasi I: lle kuvattu, ottaen huomioon, että kaksi DNA-ketjua lohkaistaan ja ei vain yksi.
Topoisomeraasi II: n aktiivisessa kohdassa proteiinin fragmentti stabiloidaan (kovalenttisen sitoutumisen avulla tyrosiiniin). kaksoiskierre DNA: ta, jota kutsutaan "fragmentiksi G". Tämä fragmentti pilkotaan ja pidetään sitoutuneena aktiiviseen kohtaan kovalenttisilla sidoksilla.
Sitten entsyymi sallii toisen DNA-fragmentin, jota kutsutaan "T-fragmentiksi", läpi fragmentin "G", joka on pilkottu entsyymin konformationaalisen muutoksen ansiosta, joka riippuu ATP: n hydrolyysistä..
Topoisomeraasi II sitoutuu "fragmentin G" kahteen päähän ja lopulta palauttaa sen alkutilan vapauttamalla fragmentin "G". Sitten DNA lieventää vääntöjännitystä, jolloin replikointi- ja transkriptioprosessit tapahtuvat.
-Ihmisen topoisomeraasit
Ihmisen genomilla on viisi topoisomeraasia: top1, top3a, top3p (tyypin I); ja top2a, top2β (tyypin II). Tärkeimmät ihmisen topoisomeraasit ovat top1 (topoisomeraasi tyyppi IB) ja 2a (topoisomeraasi tyyppi II).
Topoisomeraasin estäjät
-Topoisomeraasit kemiallisen hyökkäyksen kohteena
Koska topoisomeraasien katalysoimat prosessit ovat välttämättömiä solujen eloonjäämiseksi, nämä entsyymit ovat hyviä hyökkäyksen kohteita, jotka vaikuttavat pahanlaatuisiin soluihin. Tätä varten topoisomeraaseja pidetään tärkeinä monien ihmisten sairauksien hoidossa.
Lääkkeet, jotka ovat vuorovaikutuksessa topoisomeraasi ovat tällä hetkellä laajalti tutkittu kemoterapeuttiset aineet syöpäsoluja vastaan (eri elimissä) ja taudinaiheuttajia.
-Estotyypit
Topoisomeraasiaktiivisuuden estävät lääkkeet voivat:
- Otetaan DNA: han.
- Vaikuta topoisomeraasientsyymiin.
- Lomitetaan molekyyliin lähellä entsyymin aktiivista kohtaa, kun taas DNA-topoisomeraasikompleksi stabiloituu.
Stabilointia ohimenevä muodostama kompleksi DNA: ta sitova ja tyrosiini Katalyyttisen entsyymin estää sitoutumisen katkaistun fragmenttien, joka voi johtaa solun kuolemaan.
-Topoisomeraasien inhibiittorit
Topoisomeraaseja inhiboivia yhdisteitä ovat seuraavat.
Kasvainvastaiset antibiootit
Antibiootteja käytetään syöpää vastaan, koska ne estävät kasvainsolujen kasvua yleensä häiritsemällä niiden DNA: ta. Niitä kutsutaan usein antineoplastisiksi antibiooteiksi (syöpää vastaan). Esimerkiksi aktinomysiini D vaikuttaa topoisomeraasiin II ja sitä käytetään Wilmsin kasvaimissa lapsilla ja rabdomyosarcomissa..
antrasykliinit
Antrasykliinit ovat antibioottien joukossa tehokkaimpia syöpälääkkeitä ja laajin spektri. Niitä käytetään keuhkosyövän, munasarjojen, kohdun, mahalaukun, virtsarakon, rintojen, leukemian ja lymfoomien hoidossa. On tunnettua vaikuttaa topoisomeraasiin II interkalaatiolla DNA: ssa.
Ensimmäinen antrasykliini, joka oli eristetty aktinobakteereista (Streptomyces peucetius) oli daunorubisiini. Myöhemmin doksorubisiini syntetisoitiin laboratoriossa, ja nyt käytetään myös epirubisiinia ja idarubisiinia..
antrakinoni
Antrakinoneilla tai antraseenidionit ovat johdetut yhdisteet antraseeni samanlainen antrasykliinit, että vaikutetaan topoisomeraasi II interkalaatiolla DNA. Niitä käytetään metastasoituneen rintasyövän, non-Hodgkin-lymfooman (NHL) ja leukemia.
Näitä lääkkeitä löydettiin joidenkin hyönteisten, kasvien (frángula, senna, rabarberi), jäkälöiden ja sienien pigmenteistä; sekä hoelita, joka on luonnollinen mineraali. Annoksesta riippuen ne voivat olla syöpää aiheuttavia.
Näistä yhdisteistä meillä on mitoksantronia ja sen analogista losoksantronia. Nämä estävät pahanlaatuisten tuumorisolujen proliferaation, sitoutuvat irreversiibelisti DNA: han.
epipodofyllotoksiineja
Podofyllotoksiinit, kuten epipodofyllotoksiinit (VP-16) ja teniposidi (VM-26), muodostavat kompleksin topoisomeraasin II. Niitä käytetään vastaan keuhkosyöpä, kivessyöpä, leukemia, lymfoomat, munasarjasyövän, rintasyöpä ja pahanlaatuisia kallonsisäinen kasvaimet, mm. Ne ovat peräisin kasveista Podophyllum notatum ja P. peltatum.
Kamptotesiinianalogit
Kampotekiinit ovat yhdisteitä, jotka estävät topoisomeraasia I, ja niiden joukossa ovat irinotekaani, topotekaani ja diflomotekaani.
Näitä yhdisteitä on käytetty vastaan paksusuolen syövän, keuhko- ja rinta-, ja saadaan luonnollisesti kuoresta ja puun lehdet lajien Camptotheca acuminata kiinalaisista persikoista ja Tiibetistä.
Luonnollinen inhibitio
Topoisomeraasien I ja II rakenteelliset muutokset voivat tapahtua myös täysin luonnollisesti. Tämä voi tapahtua joidenkin tapahtumien aikana, jotka vaikuttavat sen katalyyttiseen prosessiin.
Näistä muutoksista voidaan mainita pyrimidiinidimeerien muodostuminen, typpipohjaisten emästen ja muiden oksidatiivisen stressin aiheuttamat tapahtumat..
viittaukset
- Anderson, H., & Roberge, M. (1992). DNA-topoisomeraasi II: Katsaus sen osallistumiseen kromosomirakenteeseen, DNA-replikaatioon, transkriptioon ja mitoosiin. Cell Biology International Reports, 16 (8): 717-724. doi: 10.1016 / s0309-1651 (05) 80016-5
- Chhatriwala, H., Jafri, N. & Salgia, R. (2006). Katsaus topoisomeraasin estoon keuhkosyöpään. Cancer Biology & Therapy, 5 (12): 1600 - 1607. doi: 10,4161 / cbt.5.12.3546
- Ho, Y.-P., Au-Yeung, S.C. F., ja To, K. K. W. (2003). Platinapohjaiset syöpälääkkeet: Innovatiiviset suunnittelustrategiat ja biologiset näkökulmat. Medicinal Research Reviews, 23 (5): 633-655. doi: 10.1002 / med.10038
- Li, T.-K., & Liu, L. F. (2001). Topoisomeraasi-kohdentavien lääkeaineiden aiheuttama tuumorisolukuolema. Farmakologian ja toksikologian vuosikatsaus, 41 (1): 53-77. doi: 10.1146 / annurev.pharmtox.41.1.53
- Liu, L. F. (1994). DNA-topoisomeraasit: Topoisomeraasin kohdentavat lääkkeet. Academic Press. s. 307
- Osheroff, N. ja Bjornsti, M. (2001). DNA-topoisomeraasi. Entsymologia ja lääkkeet. Vol. II. Humana Press. s. 329.
- Rothenberg, M. L. (1997). Topoisomeraasi I: n estäjät: Tarkista ja päivitä. Annals of Oncology, 8 (9), 837-855. doi: 10,1023 / a: 1008270717294
- Ryan B. (2009, joulukuu 14). Topoisomeraasi 1 ja 2. [Videotiedosto]. Haettu osoitteesta youtube.com