Jaa:



Foraminiferan ominaisuudet, morfologia, luokittelu, elinkaari
huokoseläinten jäännöksiä ne ovat ryhmä amoeboidia alkueläimiä, joidenkin meren ja muiden makean veden. He ilmestyivät alkukauden alussa (Kambrium) ja heidän seuraajansa ovat asuttaneet nykyiset valtameret. Ne löytyvät rannikkoalueilta (hypo tai hypersaliini) merenpohjaan ja trooppisista kylmiin arktisiin ja Etelämantereen valtameriin.
Sen jakautuminen riippuu useista tekijöistä. Esimerkiksi on olemassa lajeja, jotka tukevat suuria ja usein lämpötilan muutoksia, kun taas toiset eivät pysty selviytymään siitä, joten valtamerten lämpörakenne merkitsee merkittäviä eroja foraminifera-yhdistysten välillä..

Samoin syvyys on myös määrittävä tekijä foraminiferan jakautumisessa johtuen niiden suorasta esiintymisestä valon tunkeutumiseen. Toisaalta paine liittyy aikaisempiin tekijöihin (lämpötila ja syvyys), jotka vaikuttavat suoraan CO: n liukoisuuteen2, mikä vaikuttaa kalsiumkarbonaatin erittymiseen kuorien muodostumiseen.
Toisaalta matalilla alueilla veden vesi on tärkeä tekijä, koska se vaikuttaa alustan tyypiin (kovaan tai pehmeään) ja ravinteiden jakautumiseen..
Samoin muut tekijät, kuten suolapitoisuus, veden sameus, pH, hivenaineiden läsnäolo ja / tai orgaaniset komponentit, virrat, sedimentoitumisnopeus, voivat määrittää paikallisesti foraminiferaan jakautumisen..
indeksi
- 1 Tunnetut lajit
- 2 Ominaisuudet
- 2.1 Foraminiferan koon ja morfologian tekijät
- 3 Taksonomia
- 4 Luokitus
- 4.1 Athalamea
- 4.2 Monotalamea
- 4.3 Ksenofoofia
- 4.4 Tubotalamea
- 4.5 Globotalamea
- 5 Morfologia
- 5.1 -Koko
- 5.2-Protoplasma
- 5.3 -Skeleton tai kuori
- 5.4 - Foraminiferan tyypit
- 5.5 -Sudopodit
- 6 Elinkaari
- 7 Jäljentäminen
- 8 Ravitsemus
- 9 Sovellukset
- 9.1 Mauritiuksen tapaus
- 10 Viitteet
Tunnetut lajit
Tällä hetkellä tiedetään yli 10 000 lajia, ja noin 40 000 on kuollut sukupuuttoon. Joillakin lajeilla on merenpohjan elinympäristö, eli ne ovat pohjaeläimiä, jotka usein elävät naamioituneina hiekalla, joka on osa epifaaalia (epibetónicos), tai ne voivat elää hiekan alla (endobetónicos). Tästä syystä niitä kutsutaan myös eläväksi hiekaksi.
He voivat elää myös kasveissa, joissa he levittävät epifyytteinä ja jopa monet heistä valitsevat syrjäisen elämän, eli ne elävät kiinteästi substraattiin koko olemassaolonsa ajan.
Vaikka muut foraminiferat elävät kelluvat eri syvyydessä meressä (välillä 0 - 300 m), toisin sanoen ne ovat planktonisia elämiä, jotka ovat osa merimikroplanktonia. Nämä muodot ovat niukempia ja vähemmän erilaisia.
Suurin ja monimutkaisin planktoninen foraminifera ovat yleisempiä trooppisissa ja subtrooppisissa ympäristöissä. Korkealla leveysympäristössä nämä organismit ovat yleensä niukasti, pienempiä ja hyvin yksinkertaisia.
piirteet
Foraminiferassa erottuva ominaisuus on luuranko tai kuori, rakenne, joka on mahdollistanut sen, että kuolleet muodot olisi voitu tutkia meren mikrofossiilien muodossa, jotka talletetaan meren pohjalle..
Siten kuori on peruselementti foraminiferan erottamiseksi, ja se on ainoa fossiilisoituvan organismin rakenne. Nämä fossiilit ovat hyvin runsaasti meren sedimenteissä, jotka myös osallistuvat sedimenttikivien muodostumiseen.
Kuoren tärkeimmät kemialliset yhdisteet ovat kalsiitti, aragoniitti ja piidioksidi. Alkiokammion muoto ja ulottuvuus riippuvat sen alkuperästä riippumatta siitä, ovatko ne seksuaalista tai epätavallista lisääntymistä.
Ontogeniikan aikana foraminifera ohjaa kammioiden kasvua ja kokoa. Tämä ohjaus tapahtuu pseudopodiaalivirtojen pituuden ja järjestelyn kautta, koska pseudopodit ovat vastuussa orgaanisen kalvon muodostumisesta, joka edeltää mineraalikuoren.
Tämä prosessi on erittäin tärkeä soluprosessien ylläpitämiseksi, koska kamera toimii bioreaktorina.
Tekijät, jotka vaikuttavat foraminiferan kokoon ja morfologiaan
On syytä huomata, että foraminifera-aineen koko ja lopullinen morfologia riippuvat useista tekijöistä, mukaan lukien:
- Alkiokammion muoto ja mitat.
- Kasvuvaiheiden lukumäärä aikuisvaiheeseen asti (esim. Foraminifera-kammioiden lukumäärä).
- Kameran muoto ja sen muutokset ontogeneettisen aikana.
- Kamerien järjestely.
Suuremmilla foraminiferaaleilla on strategiset kasvumallit vakio- kammioiden tilavuuden ylläpitämiseksi optimaalisen koon ylittämättä. Nämä strategiat muodostuvat kameroiden jakamisesta useisiin osastoihin, joita kutsutaan cliquesiksi.
Nämä klusterit on järjestetty siten, että varmistetaan kuljetus- ja säätötoiminnot kammioiden sisäpuolella olevan protoplasmin ja ulkopuolisen välillä. Toisin sanoen kaikki kamerat ja klikkit ovat täysin toisiinsa yhteydessä.
Kammioiden järjestely voi seurata suorakulmaista tai spiraalista akselia. Tämä riippuu pseudopodiaalivirtojen sijainnista ja aukon tai aukkojen sijainnista kammiossa.
taksonomia
Alue: Eukarya
Kuningaskunta: Protista
Ilman Range: SAR Supergroup
superphylum: rhizaria
phylum: huokoseläinten jäännöksiä
Luokat ja tilaukset
- Athalamea (Reticulomyxida)
- Monothalamea (Allogromiida, Astrorhizida, Komokiida)
- Xenophyofhorea (psamminidi, stannomidi)
- Tubothalamea(fusulinida, involutinida, miliolida, silicoloculinida, spirillinida)
- Globothalamea(Lituolida, Loftusiida, Schlumbergerinida, textulariida, Trochamminida, rotaliida, buliminida, globigerinida, robertinida, Carterinida, lagenida).
luokitus
Vaikka on vielä monia asioita, joita on selvennettävä, voimme erottaa 5 luokkaa:
Athalamea
Täältä löydät foraminiferat, joilla ei ole kuoria tai jotka ovat alasti.
Monothalamea
Se sisältää betonisen foraminifera, joka esittää orgaanisen tai agglutinoidun kuoren, jossa on yksi kammio.
xenophyophores
Tässä tapauksessa foraminifera ovat erikoistuneita Betonityyppisiä, suurikokoisia, monikerroksisia ja agglutinoidun kuoren kanssa. Ne ovat yleensä detritivoreita tai saprofageja, toisin sanoen ne saavat ruokansa detriitista tai orgaanisesta aineesta hajoamassa.
Tubothalamea
Tähän kuuluu betoninen foraminifera, jolla on useita putkimaisia kammioita ainakin nuorisovaiheessa, jotka voidaan kierrättää spiraaliin sidotulla tai kalkkipitoisella kuorella.
Globothalamea
Tähän luokitukseen sisältyvät sekä betoniset että planktoniset foraminiferaat, joissa on monikamariset kuoret, joissa on globulaarinen, agglutinoitu tai kalkkipitoinen muoto. Kuoret voivat olla uniseriateja, biseriada, triseriada tai trocospiraladas.
Tämä luokitus on kuitenkin jatkuvasti kehittymässä.
morfologia
-koko
Foraminiferan koko vaihtelee yleensä välillä 0,1 - 0,5 cm, ja jotkut lajit voivat mitata 100 - 20 cm..
-protoplasma
Foraminifera muodostuu protoplasma-massasta, joka muodostaa foraminiferan solun.
Protoplasma on yleensä väritön, mutta voi joskus sisältää pieniä määriä orgaanisia pigmenttejä, lipidimateriaalia, symbioottisia leviä tai rautayhdisteitä, jotka antavat sille väriä.
Protoplasma koostuu sisäisestä osasta, jota kutsutaan endoplasmaksi ja ulkoiseksi osaksi ektoplasmaksi.
Endoplasmassa sitä suojaa kuori ja siinä organellit jakautuvat ruoansulatuskanavan vakuoleina, ydinaineina, mitokondrioina, rakeina, Golgi-laitteina tai ribosomeina. Tästä syystä sitä kutsutaan joskus rakeiseksi endoplasmaksi. Ektoplasma on läpinäkyvä ja sieltä sisäänvedettävät pseudopodit lähtevät.
Protoplasmaa ympäröi ulkoisesti orgaaninen kalvo, joka muodostuu päällystetyistä mukopolysakkaridilevyistä..
Protoplasminen massa laajenee ulos kuoresta yhden tai useamman aukon (huokoset) läpi ja peittää sen ulkopuolelta (extra-cameral protoplasm), ja näin pseudopodia muodostuu.
-Luuranko tai kuori
Foraminifera kiinnittää solupinnansa pysyvästi mineraalirunko (kuori)..
Kuori muodostuu kammioista, jotka on erotettu septa, mutta samalla ne kommunikoivat toistensa kautta ns. Luuston tai kuoren kemiallinen koostumus tekee niistä rakenteita, jotka fossiilisoivat hyvin.
Kammioiden sisätilat on peitetty orgaanisella materiaalilla, joka on hyvin samanlainen kuin kitiini. Lisäksi kuoressa voi olla pääaukot; Voit myös omistaa huokoset ulospäin tai puuttuvat niistä.
Mineraalikuori voidaan muodostaa yhdestä osastosta (primitiivinen foraminifera tai monotalamos) tai kammiosta, joka kasvaa jatkuvasti, tai useista kammioista, jotka on muodostettu peräkkäisissä vaiheissa, monimutkaisessa epäjatkuvan kasvujärjestelmässä (foraminifera polylamas).
Tämä viimeinen prosessi on lisätä aiemmin muodostettuun kuoreen ja strategisiin paikkoihin uusi luustomateriaali.
Monet foraminiferaat pystyvät valitsemaan materiaalin, joka muodostaa kuorensa kemiallisen koostumuksensa, koonsa tai muodonsa mukaisesti, koska alimmaiset pseudopodiaalivirrat, jotka ovat kosketuksissa alustan kanssa, pystyvät tunnistamaan sen.
-Foraminiferan tyypit
Kuoren rakennusmuodon mukaan ne voidaan luokitella kolmeen päätyyppiin Foraminifera:
Agglutinoidut
Tämäntyyppisessä kuoressa foraminifera kerää pseudopoodiensa kanssa suuren määrän orgaanisia aineita, jotka ovat käytettävissä ympäristössä, jossa ne elävät, ja jotka myöhemmin agglutinoivat, kuten mineraalirakeet, sienen spicules, diatomit jne..
Useimmat agglutinoidut foraminiferaat sementoivat kuorensa kalsiumkarbonaatilla, mutta jos tämä yhdiste ei ole läsnä ympäristössä, kuten esimerkiksi ne, jotka elävät meren syvillä alueilla, joissa kalsiumia ei ole, ne voivat tehdä niin piidioksidilla, ferrugiinilla, orgaanisilla sementeillä etc
porcelaneous
Tällöin kuori muodostetaan magnesiumsakkiitti- neulojen kautta, jotka syntetisoidaan foraminiferan Golgi-laitteessa.
Nämä neulat kuljetetaan ja kerätään ulkopuolelle, ja ne voivat toimia liitoselementteinä ulkomaisille rakenteille (sementti) tai suoraan muodostaa ulkoisen luuranko. Ne löytyvät hypersaliini-ympäristöissä (> 35% suolapitoisuus).
Ne ovat yleensä imperforaatteja, eli niissä on yleensä pseudo-huokoset, jotka eivät pääse kuoren läpi kokonaan.
hyaline
Nämä muodostuvat kalsiittikiteiden kasvusta orgaanisen templaatin ansiosta, joka muodostuu biomineralisointimenetelmästä (mineralisaatio in situ), joka toteutetaan ulkoisesti protoplasmaattiseen runkoon.
Niille on tunnusomaista se, että se on läpinäkyvä seinän ohuuden vuoksi. Ne on myös rei'itetty, jos huokosten sijainti, tiheys ja halkaisija vaihtelee lajin mukaan.
-seudópodos
Tätä rakennetta käytetään mobilisointiin, kiinnittymiseen substraatteihin, saalistus ja luuranko. Pseudopodien takaisinvetämiseen ja laajentamiseen foraminifera on kehittynyt mikrotubulusverkko, joka on järjestetty enemmän tai vähemmän rinnakkaisiin riveihin.
Pseudopodien pidennys voi ulottua kaksi tai kolme kertaa kehon pituudesta ja jopa jopa 20 kertaa sen pituuden. Tämä riippuu jokaisesta tietystä lajista.
Liikkumisen tyyppi siirron aikana liittyy suoraan kuoren muotoon ja aukkojen sijaintiin (missä pseudopodit ovat peräisin).
Mutta useimmat foraminifera liikkuvat seuraavasti: pseudopodit kiinnittyvät substraattiin ja työntävät sitten loput solusta. Tällä tavalla liikkuminen voi edetä nopeudella noin 1 - 2,5 cm / tunti.
Toisaalta foraminiferan pseudopoodeja kutsutaan Granurreticulopodialle, koska pseudopodien sisäpuolella on kaksisuuntainen sytoplasminen virtaus, joka kuljettaa rakeita.
Rakeet voivat koostua erilaisten materiaalien partikkeleista, mitokondrioista, ruoansulatuskanavan tai jätteen vakuoleista, synbioottisista dinoflagellateista jne. Tästä syystä yksi ryhmän synonyymeistä on Granuloreticulosa.
Toinen tärkeä pseudopodien ominaisuus on se, että ne ovat yleensä pitkiä, ohuita, haarautuneita ja hyvin runsaita ja muodostavat siten retikulopodian verkoston pinoamalla (anastomosis)..
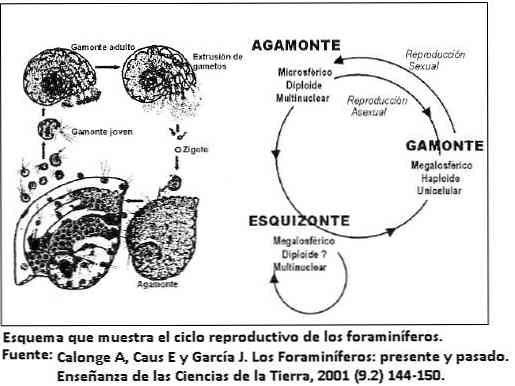
Elinkaari
Foraminiferan elinkaari on yleensä lyhyt, yleensä muutama päivä tai viikko, mutta suurissa muodoissa elinkaari voi olla kaksi vuotta.
Kesto riippuu foraminiferan hyväksymästä elämänstrategiasta. Esimerkiksi pienet muodot, joilla on yksinkertainen morfologia, kehittävät lyhyen opportunistisen strategian.
Kuitenkin suuret muodot ja kuoren poikkeuksellisen monimutkainen morfologia kehittävät konservatiivisen elämänstrategian.
Tämä viimeinen käyttäytyminen on yksirunkoisissa organismeissa hyvin harvinaista; antaa heille mahdollisuuden säilyttää yhtenäinen väestötiheys ja hidas kasvu.
kopiointi
Useimmissa foraminifera-aineissa esiintyy kahta morfologiaa, sukupolvenvaihtelu riippuen lisääntymisen tyypistä, seksuaalisesta tai aseksuaalisesta, lukuun ottamatta planktonista foraminiferaa, joka vain lisääntyy seksuaalisesti.
Tätä morfologian muutosta kutsutaan dimorfismiksi. Seksuaalisen lisääntymisen (gamogonia) syntynyttä muotoa kutsutaan gamoniksi, kun taas epätavallinen lisääntyminen (schizogony) muodostaa muodon. Molemmat ovat morfologisesti erillisiä.
Jotkut foraminiferat koordinoivat lisääntymisjaksoa kausivaiheen kanssa resurssien käytön optimoimiseksi. Ei ole harvinaista, että ennen kuin sukupuolinen sukupolvi esiintyy betonimuodoissa, esiintyy useita jatkuvia epäsuoria toistoja.
Tämä selittää, miksi kaavamuodot ovat runsaampia kuin gamontes. Gamoni esittelee aluksi yhden ytimen ja jakaa sen jälkeen tuottamaan lukuisia sukusoluja.
Vaaka on monisoluinen ja meiosisfragmenttien jälkeen muodostamassa uusia sukusoluja.
ravitsemus
Foraminiferaaleille on tunnusomaista se, että se on heterotrofinen, eli ne syöttävät orgaanista ainetta.
Tässä tapauksessa foraminifera ruokkii pääasiassa piileviä tai bakteereja, mutta muita suurempia lajeja syötetään sukkulamatoilla ja äyriäisillä. Vangit jäävät pseudopodiensa kautta.
Myös nämä organismit voivat käyttää eri tyyppisiä symbionteja, kuten vihreitä, punaisia ja kultaisia leviä, sekä piileviä ja dinoflagellateja, ja monista niistä voi olla jopa hyvin monimutkainen samassa yksilössä.
Toisaalta jotkut foraminifera-lajit ovat kleptoplastisia, mikä tarkoittaa, että nautittujen levien kloroplastit tulevat osaksi foraminiferaa jatkamaan fotosynteesitoiminnon suorittamista..
Tämä on vaihtoehtoinen tapa tuottaa energiaa elämään.
sovellukset
Foraminiferan fossiilisen levyn runsaus geologisen ajan, kehityksen, monimutkaisuuden ja koon mukaan tekee niistä suosikkivälineen tutkia maan nykyistä ja menneisyyttä (geologinen kello).
Siksi sen suuri lajien monimuotoisuus ovat erittäin hyödyllisiä biostratigrafisen, paleoekologisen, paleokeanografisen tyypin tutkimuksissa..
Se voi kuitenkin myös auttaa ehkäisemään ekologisia katastrofeja, jotka voivat vaikuttaa talouteen, koska foraminifera-populaatioiden muutokset osoittavat muutoksia ympäristössä..
Esimerkiksi foraminifera ilman kuoria, ovat herkkiä ympäristömuutoksille ja reagoivat nopeasti ympäröivään ympäristöön. Siksi ne ovat ihanteellisia indikaattorilajeja riutta-veden laadun ja terveyden tutkimuksessa.
Mauritiuksen tapaus
Myös jotkin tapahtumat ovat tehneet meistä pohdintaa tästä. Sama koskee Mauritiuksessa havaittua ilmiötä, jossa osa rannan valkoisesta hiekasta hävisi, ja nyt heidän täytyy tuoda se Madagaskarilta matkailuvirran ylläpitämiseksi.
Ja mitä siellä tapahtui? Mistä hiekka tulee? Miksi se katosi??
Vastaus on seuraava:
Hiekka ei ole vain monien organismien kalsiumkarbonaattikuorien kerääntyminen, muun muassa foraminifera, joka vedetään rannan reunaan. Hiekan katoaminen johtui karbonaattintuottajien asteittaisesta ja jatkuvasta vähenemisestä.
Tämä tapahtui merien pilaantumisen seurauksena typen ja fosforin kanssa, jotka ulottuvat rannikolle johtuen lannoitteiden liiallisesta käytöstä tiettyjen tuotteiden, kuten sokeriruoan, kylvämisessä..
Siksi on tärkeää tutkia foraminiferaa yhteiskuntatieteissä, jotta vältetään edellä kuvattujen ympäristökatastrofien, jotka vaikuttavat suoraan talouteen ja yhteiskuntaan, estäminen..
viittaukset
- Wikipedia-avustajat. huokoseläinten jäännöksiä [Online]. Wikipedia, vapaa tietosanakirja, 2018 [kuulemispäivä: 1. marraskuuta 2018]. Saatavilla osoitteessa es.wikipedia.org.
- Calonge A, Caus E ja García J. Foraminifera: läsnä ja menneisyydessä. Maantieteiden opetus, 2001 (9,2) 144-150.
- Hromic T. Biologinen monimuotoisuus ja mikrobiologi (Foraminifera: Protozoa), Boca del Guafo ja Golfo de Penas (43º-46º s), Chile. Cienc. TECNOL. 30 (1): 89 - 103, 2007
- Humphreys AF, Halfar J, Ingle JC, et ai. Meriveden lämpötilan, pH: n ja ravintoaineiden vaikutus pienen runsauden matalien veden pohjoisen foraminiferan jakautumiseen ja luonteeseen Galapagosissa. PLoS One 2018, 13 (9): e0202746. Julkaistu 2018 syyskuu 12. doi: 10.1371 / journal.pone.0202746
- De Vargas C, Norris R, Zaninetti L, Gibb SW, Pawlowski J. Molekyyliset todisteet salaisesta spekulaatiosta planktonisissa foraminifereissä ja niiden suhde valtameriin. Proc. Natl. Acad. Sei. U. A. 1999; 96 (6): 2864-8.