Jaa:



Anticodon-kuvaus, toiminnot ja ero koodonilla
antikodonista on kolmen nukleotidin sekvenssi, joka on läsnä siirto-RNA-molekyylissä (tRNA), jonka tehtävänä on tunnistaa toinen sekvenssi kolmesta nukleotidista, joka on läsnä messenger-RNA-molekyylissä (mRNA).
Tämä koodonien ja antikodonien välinen tunnistus on antiparalleeli; toisin sanoen yksi sijaitsee 5 '-> 3' suunnassa, kun taas toinen on 3 '-> 5' suunnassa. Tämä kolmen nukleotidin (tripletin) sekvenssien välinen tunnistus on olennainen käännösprosessin kannalta; toisin sanoen proteiinien synteesissä ribosomissa.
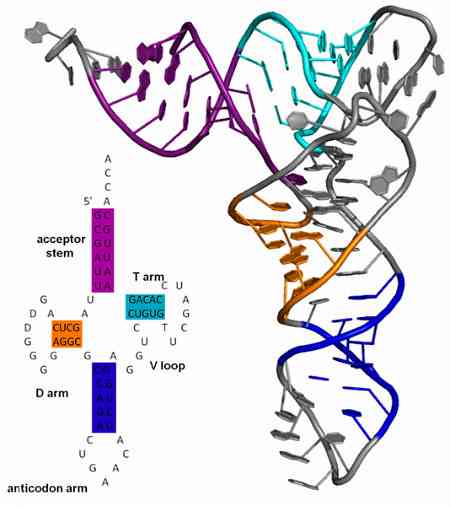
Täten siirron RNA-molekyylit "luetaan" translaation aikana niiden koodonien tunnistamisen avulla siirto-RNA: iden antikodoneilla. Nämä molekyylit kutsutaan niin, koska ne siirtävät spesifisen aminohapon proteosimolekyyliin, joka muodostuu ribosomissa.
On 20 aminohappoa, joista kukin koodaa tietty tripletti. Joitakin aminohappoja koodaa kuitenkin useampi kuin yksi tripletti.
Lisäksi jotkut kodonit tunnistavat antikodit siirto-RNA-molekyyleissä, joissa ei ole liitetty aminohappoja; nämä ovat ns. stop-kodoneja.
indeksi
- 1 Kuvaus
- 2 Toiminnot
- 3 Antikodonin ja kodonin väliset erot
- 4 Liikkumisen hypoteesi
- 4.1 RNA ja aminohapot
- 5 Viitteet
kuvaus
Antikodoni muodostuu kolmen nukleotidin sekvenssistä, jotka voivat sisältää mitä tahansa seuraavista typpipitoisista emäksistä: adeniini (A), guaniini (G), urasiili (U) tai sytosiini (C) kolmen nukleotidin yhdistelmänä siten, että se toimii kuin koodi.
Antikodonit löytyvät aina siirto-RNA-molekyyleistä ja ovat aina 3 '-> 5'-suunnassa. Näiden tRNA: iden rakenne on samanlainen kuin apila, siten, että se on jaettu neljään silmukkaan (tai silmukkaan); yhdessä silmukassa on antikodoni.
Antikodonit ovat välttämättömiä messenger-RNA: n kodonien tunnistamiseksi ja siten proteiinisynteesin prosessille kaikissa elävissä soluissa.
tehtävät
Antikodonien päätehtävä on niiden triplettien spesifinen tunnistaminen, jotka muodostavat koodonit messenger-RNA-molekyyleissä. Nämä koodonit ovat ohjeita, jotka on kopioitu DNA-molekyylistä proteiinin aminohappojen järjestyksen sanelemiseksi.
Koska transkriptio (messenger-RNA-kopioiden synteesi) tapahtuu 5 '-> 3' -suunnassa, niin messenger-RNA: n koodonit ovat tämän suunnan mukaisia. Siksi siirto-RNA-molekyyleissä olevien antikodonien on oltava vastakkaiseen suuntaan, 3 '-> 5'.
Tämä liitto johtuu täydentävyydestä. Jos esimerkiksi yksi kodoni on 5'-AGG-3 ', antikodoni on 3'-UCC-5'. Tämän tyyppinen spesifinen vuorovaikutus kodonien ja antikodonien välillä on tärkeä vaihe, joka sallii messenger-RNA: n nukleotidisekvenssin koodaamaan aminohapposekvenssin proteiinissa..
Antikodonin ja kodonin väliset erot
- Antikodonit ovat tRNA: iden trinukleotidiyksiköitä, jotka ovat komplementaarisia mRNA: iden kodoneille. Ne mahdollistavat tRNA: iden tuottaa oikeat aminohapot proteiinin tuotannon aikana. Sen sijaan kodonit ovat trinukleotidien yksiköitä DNA: ssa tai mRNA: ssa, jotka koodaavat spesifistä aminohappoa proteiinisynteesissä.
- Antikodonit ovat yhteys mRNA: n nukleotidisekvenssin ja proteiinin aminohapposekvenssin välillä. Sitä vastoin kodonit siirtävät geneettistä informaatiota ytimestä, jossa DNA on ribosomeihin, joissa tapahtuu proteiinisynteesi..
- Antikodoni löytyy tRNA-molekyylin antikodonivarresta, toisin kuin koodonit, jotka sijaitsevat DNA- ja mRNA-molekyylissä.
- Antikodoni on komplementaarinen vastaavan kodonin kanssa. Sitä vastoin mRNA: n kodoni on komplementaarinen DNA: n tietyn geenin nukleotidien tripletille..
- TRNA sisältää antikodonia. Sitä vastoin mRNA sisältää useita kodoneja.
Liukuva hypoteesi
Tasapainotushypoteesi ehdottaa, että messenger-RNA: n kodonin kolmannen nukleotidin ja siirto-RNA: n antikodonin ensimmäisen nukleotidin väliset liitokset ovat vähemmän spesifisiä kuin kahden toisen nukleotidin väliset liitokset.
Crick kuvaili tätä ilmiötä jokaisen kodonin kolmannessa sijainnissa. Tässä tilanteessa tapahtuu jotakin, joka sallii ammattiliittojen olla tavallista vähemmän tiukkoja. Se tunnetaan myös nimellä wobbling tai tamboleo.
Tämä Crick-vaimennushypoteesi selittää, kuinka tietyn tRNA: n antikodoni voidaan yhdistää kahden tai kolmen eri mRNA-kodonin kanssa.
Crick ehdotti, että koska emäsparin (tRNA: n antikodonin pohjan 59 ja mRNA: n kodonin kodin 39) välillä on vähemmän tiukkaa kuin normaali, tässä sivustossa sallitaan tietty "heiluminen" tai vähentynyt affiniteetti..
Tämän seurauksena yksittäinen tRNA tunnistaa usein kaksi tai kolme vastaavista kodoneista, jotka määrittävät tietyn aminohapon.
Normaalisti, tRNA-antikodonien emästen ja mRNA-kodonien väliset vety- sidokset noudattavat tiukkoja peruspoistussääntöjä vain kodonin kahdelle ensimmäiselle emäkselle. Tämä vaikutus ei kuitenkaan esiinny kaikkien mRNA-kodonien kaikissa kolmansissa asennoissa.
RNA ja aminohapot
Heilahdushypoteesin perusteella ennustettiin ainakin kahden siirto-RNA: n olemassaoloa kullekin aminohapolle koodoneilla, joilla oli täydellinen degeneraatio, mikä on osoittautunut totta.
Tämä hypoteesi ennusti myös kolmen siirto-RNA: n esiintymisen kaikille kuudelle seriinikodonille. Kolme tRNA: ta seriinille on todettu:
- TRNA seriinille 1 (anticodon AGG) sitoutuu koodoneihin UCU ja UCC.
- TRNA seriinille 2 (antikodon AGU) sitoutuu koodoneihin UCA ja UCG.
- TRNA seriinille 3 (anticodon UCG) sitoutuu koodoneihin AGU ja AGC.
Nämä spesifisyydet todistettiin puhdistettujen amino- asyyli-tRNA-trinukleotidien stimuloidulla sitoutumisella ribosomeihin in vitro.
Lopuksi useat siirto-RNA: t sisältävät inosiiniemäksen, joka on valmistettu hypoksantiini-puriinista. Inosiini tuotetaan adenosiinin transkription jälkeisellä modifikaatiolla.
Crick-vaimennushypoteesi ennusti, että kun inosiini on läsnä antikodonin 5'-päässä (värähtelyasento), se muodostaisi parin urasiilin, sytosiinin tai adeniinin kanssa kodonissa.
Itse asiassa puhdistettu alanyyli-tRNA, joka sisältää inosiinia (I) antikodonin 5'-asemassa, sitoutuu ribosomeihin, jotka on aktivoitu GCU: n, GCC: n tai GCA: n trinukleotideilla.
Sama tulos on saatu muilla tRNA: lla, jotka on puhdistettu inosiinilla antikodonin 5'-asennossa. Siksi Crickin heilahdushypoteesi selittää erittäin hyvin tRNA: iden ja kodonien väliset suhteet, joilla on geneettinen koodi, joka on rappeutunut, mutta tilattu.
viittaukset
- Brooker, R. (2012). Geneettiset käsitteet (Ensimmäinen toim.). McGraw-Hill-yhtiöt, Inc.
- Brown, T. (2006). Genomit 3 (3rd). Garland Science.
- Griffiths, A., Wessler, S., Carroll, S. & Doebley, J. (2015). Johdatus geneettiseen analyysiin (11. painos). W.H. Freeman
- Lewis, R. (2015). Ihmisen geenit: käsitteet ja sovellukset(11. painos). McGraw-Hill koulutus.
- Snustad, D. & Simmons, M. (2011). Geneettiset periaatteet(6. painos). John Wiley ja Sons.