Jaa:



Pentoosifaasien ja niihin liittyvien sairauksien polut
pentoosien polku Fosfaatti, joka tunnetaan myös heksoosimonofosfaatin poikkeamana, on perustavanlaatuinen metabolinen reitti, jolla on lopullisena tuotteena ribosomeja, joita tarvitaan nukleotidi- ja nukleiinihapposynteesireiteille, kuten DNA, RNA, ATP, NADH, FAD ja koentsyymi A.
Se tuottaa myös NADPH: ta (nikotiiniamidideniinidinukleotidifosfaatti), jota käytetään erilaisissa entsymaattisissa reaktioissa. Tämä reitti on hyvin dynaaminen ja kykenee mukauttamaan tuotteitaan solujen hetkellisten tarpeiden mukaan.
ATP: tä (adenosiinitrifosfaattia) pidetään solun "energiavaluuttana", koska sen hydrolyysi voidaan kytkeä moniin biokemiallisiin reaktioihin.
Samalla tavalla NADPH on toinen välttämätön energiavaluutta rasvahappojen pelkistävälle synteesille, kolesterolin synteesille, neurotransmitterisynteesille, fotosynteesille ja vieroitusreaktioille..
Vaikka NADPH ja NADH ovat rakenteeltaan samanlaisia, niitä ei voida käyttää vaihtelevasti biokemiallisissa reaktioissa. NADPH osallistuu vapaan energian käyttöön tiettyjen metaboliittien hapetuksessa pelkistäväksi biosynteesiksi.
Sitä vastoin NADH osallistuu vapaan energian käyttöön metaboliittien hapetuksesta ATP: n syntetisoimiseksi.
indeksi
- 1 Historia ja sijainti
- 2 Toiminnot
- 3 vaihetta
- 3.1 Hapetusvaihe
- 3.2 Ei-hapettava vaihe
- 4 Liittyvät sairaudet
- 5 Viitteet
Historia ja sijainti
Merkkejä tämän reitin olemassaolosta alkoi vuonna 1930 kiitos tutkija Otto Warburg, jolle NADP: n löytäminen johtuu hänelle+.
Tietyt havainnot mahdollistivat reitin löytämisen, erityisesti hengityksen jatkumisen glykolyysin estäjien, kuten fluoridi-ionin, läsnä ollessa..
Sitten 1950 tutkijat Frank Dickens, Bernard Horecker, Fritz Lipmann ja Efraim Racker kuvailivat pentoosifosfaattireittiä.
Kolesterolin ja rasvahappojen synteesiin osallistuvien kudosten, kuten rintarauhasen, rasvakudoksen ja munuaisten, pitoisuudet ovat suuria pentoosifosfaattientsyymejä.
Maksa on myös tärkeä kudos tälle reitille: noin 30% glukoosin hapetuksesta tässä kudoksessa tapahtuu pentoosifosfaattireitin entsyymien ansiosta..
tehtävät
Pentoosifosfaattireitti vastaa hiilen homeostaasin ylläpidosta solussa. Samoin polku syntetisoi aminohappojen synteesiin osallistuvien nukleotidien ja molekyylien prekursorit (peptidien ja proteiinien rakenteelliset lohkot).
Se on tärkein entsymaattisten reaktioiden vähentämisen teho. Lisäksi se tarjoaa tarvittavat molekyylit anabolisille reaktioille ja puolustusprosesseille oksidatiivista stressiä vastaan. Kappaleen viimeinen vaihe on kriittinen redox-prosesseissa stressitilanteissa.
faasit
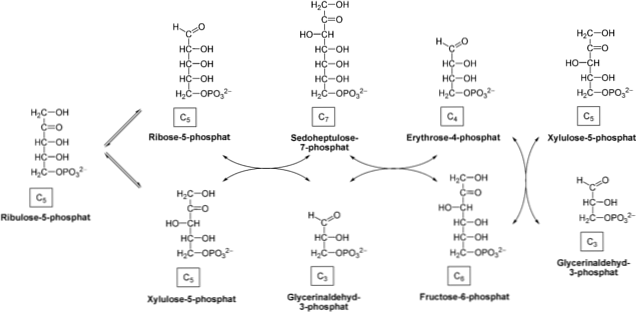
Pentoosifosfaattireitti koostuu kahdesta faasista sellulaarisessa sytosolissa: hapettava, joka tuottaa NADPH: n hapettamalla glukoosi-6-fosfaatti riboos-5-fosfaatiksi; ja ei-hapettava, joka käsittää kolmen, neljän, viiden, kuuden ja seitsemän hiilen sokerien muuntamisen.
Tällä reitillä on yhteisiä reaktioita Calvin-syklin ja Entner-Doudoroff-reitin kanssa, joka on vaihtoehto glykolyysille.
Oksidatiivinen vaihe
Hapetusvaihe alkaa glukoosi-6-fosfaatti-molekyylin dehydrogenaatiosta hiilellä 1. Tämä reaktio katalysoi glukoosi-6-fosfaatti-dehydrogenaasin entsyymi, jolla on NADP: n korkea spesifisyys.+.
Tämän reaktion tuote on 6-fosfonoglukono-5-laktoni. Tämän jälkeen entsyymi laktonaasi hydrolysoi tämän tuotteen, jolloin saadaan 6-fosfoglukonaatti. Jälkimmäinen yhdiste otetaan 6-fosfoglukonaatti-dehydrogenaasin entsyymillä ja siitä tulee ribuloosin 5-fosfaatti.
Fosfenttoosi-isomeraasi entsyymi katalysoi oksidatiivisen faasin viimeistä vaihetta, joka käsittää riboosin 5-fosfaatin synteesin ribotoosin 5-fosfaatin isomeroinnilla.
Tämä reaktiosarja tuottaa kaksi NADPH-molekyyliä ja yksi ribose-5-fosfaatin molekyyli glukoosi-6-fosfaatin molekyyliä kohti, joka siirtyy tähän entsymaattiseen reittiin.
Joissakin soluissa NADPH-vaatimukset ovat suuremmat kuin ribose-5-fosfaatin vaatimukset. Siksi transketolaasi- ja transaldolaasientsyymit ottavat riboosin 5-fosfaatin ja muuttavat sen glyseraldehydi-3-fosfaatiksi ja fruktoosi-6-fosfaatiksi, jolloin saadaan aikaan ei-hapettava faasi. Nämä kaksi viimeistä yhdistettä voivat tulla glykolyyttiseen reittiin.

Ei-hapettava faasi
Faasi alkaa epimerointireaktiolla, jota katalysoi entsyymi pentoosi-5-fosfaattiepimeraasi. Tämä entsyymi ottaa ribuloos-5-fosfaatin ja muunnetaan ksyluloosi-5-fosfaatiksi.
Transketolaasientsyymi, joka toimii yhdessä koentsyymitiamiinipyrofosfaatin (TTP) kanssa, siirtyy tuotteeseen, joka katalysoi ksyluloos-5-fosfaatin kulkeutumista riboos-5-fosfaattiin. Ketoosin siirrosta aldoosiin valmistetaan glyseraldehydi-3-fosfaatti ja sedohepuloos-7-fosfaatti.
Seuraavaksi transaldolaasientsyymi siirtää C3: n sedoheptuloosi-7-fosfaatti-molekyylistä glyseraldehydi-3-fosfaattiin, joka tuottaa neljän hiilen sokerin (erytroosi-4-fosfaatin) ja kuuden hiilen sokerin (fruktoosi-6). fosfaattia). Nämä tuotteet pystyvät syöttämään glykolyyttisen reitin.
Transketosala-entsyymi vaikuttaa jälleen siirtämään ksyluloos-5-fosfaatin C2 erytroosin-4-fosfaattiin, jolloin saadaan fruktoosi-6-fosfaatti ja glyseraldehydi-3-fosfaatti. Kuten edellisessä vaiheessa, nämä tuotteet voivat päästä glykolyysiin.
Tämä toinen vaihe yhdistää NADPH: a tuottavat reitit ATP: n ja NADH: n syntetisoinnista vastaaviin. Lisäksi fruktoosi-6-fosfaatti ja glyseraldehydi-3-fosfaatti voivat päästä gluko- geneesiin.
Liittyvät sairaudet
Eri patologiat liittyvät pentoosifosfaatin kulkuun, näiden neuromuskulaaristen sairauksien ja eri syöpätyyppien välillä.
Useimmat kliiniset tutkimukset keskittyvät glukoosi-6-fosfaattidehydrogenaasin aktiivisuuden kvantifiointiin, koska se on tärkein entsyymi, joka vastaa reitin säätelystä.
Anemiaan alttiisiin yksilöihin kuuluvissa verisoluissa heillä on alhainen glukoosi-6-fosfaattihydrogenaasin entsymaattinen aktiivisuus. Sitä vastoin karsinoomiin liittyvät solulinjat kurkunpäässä osoittavat korkeaa entsyymiaktiivisuutta.
NADPH osallistuu glutationin tuotantoon, joka on keskeinen peptidimolekyyli, joka suojaa hapettumisreaktioita hapettuvassa stressissä..
Erilaiset syöpätyypit johtavat pentoosireitin aktivoitumiseen ja liittyvät metastaasiin, angiogeneesiin ja vasteisiin kemoterapiaan ja sädehoitoon..
Toisaalta krooninen granulomatoottinen sairaus kehittyy, kun NADPH: n tuotannossa on puutetta.
viittaukset
- Berg, J.M., Tymoczko, J.L., Stryer, L (2002). biokemia. WH Freeman
- Konagaya, M., Konagaya, Y., Horikawa, H., & Iida, M. (1990). Pentoosifosfaattireitti neuromuskulaarisissa sairauksissa - lihasglukoosin 6-fosfaattihydrogenaasiaktiivisuuden ja RNA-pitoisuuden arviointi. Rinsho shinkeigak. Kliininen neurologia, 30(10), 1078-1083.
- Kowalik, M.A., Columbano, A., & Perra, A. (2017). Pentoosifosfaattireitin kehittyvä rooli hepatosellulaarisessa karsinoomassa. Onkologian rajat, 7, 87.
- Patra, K. C., & Hay, N. (2014). Pentoosifosfaattireitti ja syöpä. Biokemiallisten tieteiden suuntaukset, 39(8), 347 - 354.
- Stincone, A., Prigione, A., Cramer, T., Wamelink, M., Campbell, K., Cheung, E., ... & Keller, M. A. (2015). Metabolian paluu: pentoosifosfaattireitin biokemia ja fysiologia. Biologiset arviot, 90(3), 927-963.
- Voet, D., & Voet, J. G. (2013). biokemia. Artmed Publisher.